Los antipiréticos para niños los prescribe un pediatra. Pero hay situaciones de emergencia con fiebre en las que es necesario administrar medicamentos al niño de inmediato. Entonces los padres asumen la responsabilidad y utilizan fármacos antipiréticos. ¿Qué se le permite dar a los bebés? ¿Cómo se puede bajar la temperatura en niños mayores? ¿Qué medicamentos son los más seguros?
Material de Wikipedia: la enciclopedia libre
cuadrícula de Punnett, o rejilla pannet, - una tabla propuesta por el genetista inglés Reginald Punnett (1875-1967) como una herramienta que es un registro gráfico para determinar la compatibilidad de alelos de genotipos parentales. A lo largo de un lado del cuadrado se encuentran los gametos femeninos y, en el otro, los masculinos. Esto hace que sea más fácil y visual presentar los genotipos obtenidos mediante el cruce de gametos parentales.
Cruz monohíbrida
En este ejemplo, ambos organismos tienen el genotipo Bb. Pueden producir gametos que contienen el alelo B o b (el primero significa dominancia, el segundo significa recesivo). La probabilidad de tener un descendiente con el genotipo BB es del 25%, Bb - 50%, bb - 25%.
| Materno | |||
|---|---|---|---|
| B | b | ||
| Paternal | B | CAMA Y DESAYUNO | Cama y desayuno |
| b | Cama y desayuno | cama y desayuno | |
Los fenotipos se obtienen en una combinación 3:1. Un ejemplo clásico es el color del pelaje de una rata: por ejemplo, B - pelaje negro, b - blanco. En este caso, el 75% de la descendencia tendrá pelaje negro (BB o Bb), mientras que sólo el 25% tendrá pelaje blanco (bb).
Cruz dihíbrida
El siguiente ejemplo ilustra un cruce dihíbrido entre plantas de guisantes heterocigotas. A representa el alelo dominante de forma (guisantes redondos), a representa el alelo recesivo (guisantes arrugados). B representa el alelo dominante de color (guisantes amarillos), b es el alelo recesivo (verde). Si cada planta tiene el genotipo AaBb, entonces, dado que los alelos de forma y color son independientes, puede haber cuatro tipos de gametos en todas las combinaciones posibles: AB, Ab, aB y ab.
| AB | ab | AB | ab | |
|---|---|---|---|---|
| AB | AABB | AABb | AaBB | AaBb |
| ab | AABb | AAbb | AaBb | Aabb |
| AB | AaBB | AaBb | aaBB | aaBb |
| ab | AaBb | Aabb | aaBb | aabb |
Rinde 9 guisantes amarillos redondos, 3 guisantes verdes redondos, 3 guisantes amarillos arrugados, 1 guisante verde arrugado. Los fenotipos en un cruce dihíbrido se combinan en una proporción de 9:3:3:1.
método de árbol
Existe un método alternativo basado en árboles, pero no muestra los genotipos de gametos correctamente:
Es beneficioso utilizarlo al cruzar.
explorador checo Gregor Mendel(1822-1884) considerado fundador de la genética, ya que fue el primero, incluso antes de que tomara forma esta ciencia, en formular las leyes básicas de la herencia. Muchos científicos antes que Mendel, incluido el destacado hibridador alemán del siglo XVIII. I. Kelreuter señaló que al cruzar plantas pertenecientes a diferentes variedades, se observa una gran variabilidad en la descendencia híbrida. Sin embargo, nadie pudo explicar la compleja división y, además, reducirla a fórmulas precisas debido a la falta de un método científico de análisis hibridológico.
Fue gracias al desarrollo del método hibridológico que Mendel logró evitar las dificultades que habían confundido a los investigadores anteriores. G. Mendel informó sobre los resultados de su trabajo en 1865 en una reunión de la Sociedad de Científicos Naturales en Brünn. El trabajo en sí, titulado "Experimentos sobre híbridos de plantas", se publicó más tarde en las "Proceedings" de esta sociedad, pero no recibió la evaluación adecuada de sus contemporáneos y permaneció olvidado durante 35 años.
Como monje, G. Mendel llevó a cabo sus experimentos clásicos cruzando diferentes variedades de guisantes en el jardín del monasterio de Brünn. Seleccionó 22 variedades de guisantes que tenían claras diferencias alternativas en siete características: semillas amarillas y verdes, lisas y arrugadas, flores rojas y blancas, plantas altas y bajas, etc. Una condición importante del método hibridológico era el uso obligatorio de puros, es decir, como padres. formas que no se dividen según las características estudiadas.
La elección acertada del objeto jugó un papel importante en el éxito de la investigación de Mendel. Los guisantes son autopolinizadores. Para obtener híbridos de primera generación, Mendel castró las flores de la planta madre (eliminó las anteras) y polinizó artificialmente los pistilos con el polen del progenitor masculino. Al obtener híbridos de segunda generación, este procedimiento ya no era necesario: simplemente dejó que los híbridos F 1 se autopolinizaran, lo que hizo que el experimento requiriera menos mano de obra. Las plantas de guisantes se reprodujeron exclusivamente sexualmente, por lo que ninguna desviación podría distorsionar los resultados del experimento. Y finalmente, en los guisantes, Mendel descubrió un número suficiente de pares de caracteres brillantemente contrastantes (alternativos) y fácilmente distinguibles para el análisis.
Mendel comenzó su análisis con el tipo de cruce más simple: el monohíbrido, en el que los padres difieren en un par de rasgos. El primer patrón de herencia descubierto por Mendel fue que todos los híbridos de primera generación tenían el mismo fenotipo y heredaban el rasgo de uno de los padres. Mendel llamó a este rasgo dominante. Un rasgo alternativo del otro progenitor, que no aparecía en los híbridos, se denominaba recesivo. El patrón descubierto fue nombrado I de la ley de Mendel, o ley de uniformidad de híbridos de 1ª generación.. Durante el análisis de la segunda generación, se estableció un segundo patrón: la división de los híbridos en dos clases fenotípicas (con un rasgo dominante y con un rasgo recesivo) en determinadas proporciones numéricas. Al contar el número de individuos en cada clase fenotípica, Mendel estableció que la división en un cruce monohíbrido corresponde a la fórmula 3: 1 (tres plantas con un rasgo dominante, una con un rasgo recesivo). Este patrón se llama Ley de Mendel II, o ley de segregación. En el análisis de los siete pares de características surgieron patrones abiertos, a partir de los cuales el autor llegó a la conclusión de su universalidad. Al autopolinizar híbridos F 2, Mendel obtuvo los siguientes resultados. Las plantas con flores blancas produjeron descendencia sólo con flores blancas. Las plantas con flores rojas se comportaron de manera diferente. Sólo un tercio de ellos dio descendencia uniforme con flores rojas. La descendencia del resto se dividió en una proporción de colores rojo y blanco en una proporción de 3: 1.
A continuación se muestra un diagrama de la herencia del color de la flor de los guisantes, que ilustra las leyes I y II de Mendel.
En un intento de explicar la base citológica de los patrones abiertos, Mendel formuló la idea de inclinaciones hereditarias discretas contenidas en los gametos y que determinan el desarrollo de caracteres alternativos emparejados. Cada gameto lleva un depósito hereditario, es decir. es “puro”. Tras la fecundación, el cigoto recibe dos depósitos hereditarios (uno de la madre, otro del padre), que no se mezclan y posteriormente, cuando los gametos son formados por el híbrido, acaban también en gametos diferentes. Esta hipótesis de Mendel se denominó regla de “pureza de gametos”. La combinación de inclinaciones hereditarias en el cigoto determina el carácter que tendrá el híbrido. Mendel denotó con mayúscula la inclinación que determina el desarrollo de un rasgo dominante ( A), y recesivo se capitaliza ( A). Combinación Automóvil club británico Y Ah en el cigoto determina el desarrollo de un rasgo dominante en el híbrido. Un rasgo recesivo aparece sólo cuando se combina ah.
En 1902, V. Betson propuso designar el fenómeno de los caracteres emparejados con el término "alelomorfismo" y, en consecuencia, los caracteres mismos, "alelomorfos". Según su propuesta, los organismos que contienen las mismas inclinaciones hereditarias comenzaron a llamarse homocigotos, y aquellos que contienen diferentes inclinaciones, heterocigotos. Más tarde, el término "alelomorfismo" fue reemplazado por el término más corto "alelismo" (Johansen, 1926), y las inclinaciones hereditarias (genes) responsables del desarrollo de rasgos alternativos se denominaron "alélicas".
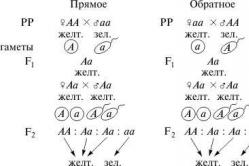
El análisis hibridológico implica el cruce recíproco de formas parentales, es decir. utilizando el mismo individuo primero como padre materno (cruce directo) y luego como padre paterno (retrocruzamiento). Si ambos cruces producen los mismos resultados, correspondientes a las leyes de Mendel, entonces esto indica que el rasgo analizado está determinado por un gen autosómico. De lo contrario, el rasgo está vinculado al sexo, debido a la localización del gen en el cromosoma sexual.

Designaciones de letras: P - individuo parental, F - individuo híbrido, ♀ y ♂ - individuo femenino o masculino (o gameto),
La letra mayúscula (A) es una disposición hereditaria dominante (gen), la letra minúscula (a) es un gen recesivo.
Entre los híbridos de segunda generación con semillas de color amarillo hay tanto homocigotos como heterocigotos dominantes. Para determinar el genotipo específico de un híbrido, Mendel propuso cruzar el híbrido con una forma homocigótica recesiva. Se llama analizar. Al cruzar un heterocigoto ( Ah) con la línea del analizador (aa), se observa la división tanto por genotipo como por fenotipo en una proporción de 1:1.

Si uno de los padres es una forma homocigótica recesiva, entonces el cruce analizador se convierte simultáneamente en un retrocruzamiento, un cruce de retorno del híbrido con la forma parental. Los descendientes de tal cruz son designados Pensión completa.
Los patrones que Mendel descubrió en su análisis de cruces monohíbridos también aparecieron en cruces dihíbridos en los que los padres diferían en dos pares de rasgos alternativos (por ejemplo, color de semilla amarillo y verde, forma lisa y arrugada). Sin embargo, el número de clases fenotípicas en F 2 se duplicó y la fórmula de división fenotípica fue 9: 3: 3: 1 (para 9 individuos con dos rasgos dominantes, tres individuos cada uno con un rasgo dominante y uno recesivo, y un individuo con dos rasgos recesivos).
Para facilitar el análisis de la división en F 2, el genetista inglés R. Punnett propuso una representación gráfica de la misma en forma de celosía, que comenzó a llevar su nombre ( cuadrícula de Punnett). A la izquierda, verticalmente, contiene los gametos femeninos del híbrido F1, y a la derecha, los masculinos. Los cuadrados interiores de la red contienen las combinaciones de genes que surgen cuando se fusionan y el fenotipo correspondiente a cada genotipo. Si los gametos se colocan en una red en la secuencia que se muestra en el diagrama, entonces en la red se puede notar el orden en la disposición de los genotipos: todos los homocigotos están ubicados a lo largo de una diagonal y los heterocigotos para dos genes (diheterocigotos) están ubicados a lo largo el otro. Todas las demás células están ocupadas por monoheterocigotos (heterocigotos para un gen).
La escisión en F 2 se puede representar mediante radicales fenotípicos, es decir indicando no el genotipo completo, sino sólo los genes que determinan el fenotipo. Esta entrada se ve así:

Los guiones en los radicales significan que los segundos genes alélicos pueden ser dominantes o recesivos, y el fenotipo será el mismo.
Esquema de cruce dihíbrido
(Cuadrícula de canastilla)

| AB | ab | AB | ab | |
| AB | AABB amarillo Cap. |
AABb amarillo Cap. |
AaBB amarillo Cap. |
AaBb amarillo Cap. |
| ab | AABb amarillo Cap. |
AAbb amarillo arruga |
AaBb amarillo Cap. |
Aabb amarillo arruga |
| AB | AaBB amarillo Cap. |
AaBb amarillo Cap. |
aaBB verde Cap. |
aaBb verde Cap. |
| ab | AaBb amarillo Cap. |
Aabb amarillo arruga |
aaBb verde Cap. |
aabb |
El número total de genotipos F2 en la red de Punnett es 16, pero hay 9 diferentes, ya que algunos genotipos se repiten. La frecuencia de diferentes genotipos se describe mediante la regla:
En un cruce dihíbrido F2, todos los homocigotos ocurren una vez, los monoheterocigotos dos veces y los diheterocigotos cuatro veces. La cuadrícula de Punnett contiene 4 homocigotos, 8 monoheterocigotos y 4 diheterocigotos.
La segregación por genotipo corresponde a la siguiente fórmula:
1AABB: 2AABBb: 1AAbb: 2AaBB: 4AaBBb: 2Aabb: 1aaBB: 2aaBBb: 1aabb.
Abreviado como 1:2:1:2:4:2:1:2:1.
Entre los híbridos F 2, solo dos genotipos repiten los genotipos de las formas parentales: AABB Y aabb; en el resto se produjo recombinación de genes parentales. Condujo al surgimiento de dos nuevas clases fenotípicas: semillas arrugadas amarillas y semillas lisas verdes.
Habiendo analizado los resultados del cruce dihíbrido para cada par de caracteres por separado, Mendel estableció el tercer patrón: la naturaleza independiente de la herencia de diferentes pares de caracteres ( Ley de Mendel III). La independencia se expresa en el hecho de que la división para cada par de características corresponde a la fórmula del cruce monohíbrido 3: 1. Por lo tanto, un cruce dihíbrido se puede representar como dos monohíbridos que ocurren simultáneamente.
Como se estableció más tarde, el tipo de herencia independiente se debe a la localización de genes en diferentes pares de cromosomas homólogos. La base citológica de la segregación mendeliana es el comportamiento de los cromosomas durante la división celular y la posterior fusión de los gametos durante la fertilización. En la profase I de la división reductora de la meiosis, los cromosomas homólogos se conjugan y luego en la anafase I divergen hacia diferentes polos, por lo que los genes alélicos no pueden ingresar al mismo gameto. Cuando divergen, los cromosomas no homólogos se combinan libremente entre sí y se mueven hacia los polos en diferentes combinaciones. Esto determina la heterogeneidad genética de las células germinales y, después de su fusión durante el proceso de fertilización, la heterogeneidad genética de los cigotos y, como consecuencia, la diversidad genotípica y fenotípica de la descendencia.
La herencia independiente de diferentes pares de rasgos facilita el cálculo de fórmulas de segregación en cruces di y polihíbridos, ya que se basan en fórmulas de cruces monohíbridos simples. Al calcular, se utiliza la ley de probabilidad (la probabilidad de que ocurran dos o más fenómenos al mismo tiempo es igual al producto de sus probabilidades). Un cruce dihíbrido se puede descomponer en dos y un cruce trihíbrido en tres cruces monohíbridos independientes, en cada uno de los cuales la probabilidad de manifestación de dos rasgos diferentes en F 2 es igual a 3: 1. Por lo tanto, la fórmula para dividir el fenotipo en el cruce dihíbrido F 2 será:
(3: 1) 2 = 9: 3: 3: 1,
trihíbrido (3:1) 3 = 27:9:9:9:3:3:3:1, etc.
El número de fenotipos en un cruce polihíbrido F2 es igual a 2 n, donde n es el número de pares de características en las que difieren los individuos progenitores.
Las fórmulas para calcular otras características de los híbridos se presentan en la Tabla 1.
Tabla 1. Patrones cuantitativos de segregación en crías híbridas
para varios tipos de cruces
| Características cuantitativas | tipo de cruce | ||
| monohíbrido | dihíbrido | polihíbrido | |
| Número de tipos de gametos formados por el híbrido F 1 | 2 | 2 2 | 2n |
| Número de combinaciones de gametos durante la formación de F 2 | 4 | 4 2 | 4n |
| Número de fenotipos F 2 | 2 | 2 2 | 2n |
| Número de genotipos F 2 | 3 | 3 2 | 3 |
|
División del fenotipo en F 2. |
3: 1 | (3: 1) 2 | (3:1)n |
| Segregación por genotipo en F 2 | 1: 2: 1 | (1: 2: 1) 2 | (1:2:1)n |
La manifestación de los patrones de herencia descubiertos por Mendel sólo es posible bajo determinadas condiciones (independientes del experimentador). Ellos son:
- Formación igualmente probable por hibridomas de todas las variedades de gametos.
- Todas las combinaciones posibles de gametos durante el proceso de fecundación.
- Igual viabilidad de todas las variedades de cigotos.
Si no se cumplen estas condiciones, entonces cambia la naturaleza de la segregación en la descendencia híbrida.
La primera condición puede violarse debido a la inviabilidad de uno u otro tipo de gameto, posiblemente por diversas razones, por ejemplo, el efecto negativo de otro gen manifestado a nivel gamético.
La segunda condición se viola en el caso de la fecundación selectiva, en la que se produce una fusión preferencial de determinados tipos de gametos. Además, un gameto con el mismo gen puede comportarse de manera diferente durante el proceso de fecundación, dependiendo de si es femenino o masculino.
La tercera condición suele violarse si el gen dominante tiene un efecto letal en el estado homocigoto. En este caso, en el cruce monohíbrido F 2 como resultado de la muerte de homocigotos dominantes Automóvil club británico en lugar de una división de 3:1, se observa una división de 2:1. Ejemplos de estos genes son: el gen del color de pelaje platino en los zorros, el gen del color de pelaje gris en las ovejas Shirazi. (Más detalles en la próxima conferencia).
El motivo de la desviación de las fórmulas de segregación mendeliana también puede ser una manifestación incompleta del rasgo. El grado de manifestación de la acción de los genes en el fenotipo se indica con el término expresividad. Para algunos genes es inestable y altamente dependiente de condiciones externas. Un ejemplo es el gen recesivo del color negro del cuerpo en Drosophila (mutación ébano), cuya expresividad depende de la temperatura, por lo que los individuos heterocigotos para este gen pueden tener un color oscuro.
El descubrimiento de Mendel de las leyes de la herencia se adelantó más de tres décadas al desarrollo de la genética. La obra "Experiencia con híbridos de plantas" publicada por el autor no fue comprendida ni apreciada por sus contemporáneos, incluido Charles Darwin. La razón principal es que en el momento de la publicación del trabajo de Mendel aún no se habían descubierto los cromosomas y aún no se había descrito el proceso de división celular que, como se mencionó anteriormente, constituía la base citológica de los patrones mendelianos. Además, el propio Mendel dudaba de la universalidad de los patrones descubiertos cuando, siguiendo el consejo de K. Nägeli, comenzó a comprobar los resultados obtenidos en otro objeto: la vellosilla. Sin saber que la tortuga carey se reproduce partenogenéticamente y, por tanto, es imposible obtener híbridos de ella, Mendel quedó completamente desanimado por los resultados de los experimentos, que no encajaban en el marco de sus leyes. Bajo la influencia del fracaso, abandonó su investigación.
El reconocimiento llegó a Mendel a principios del siglo XX, cuando en 1900 tres investigadores, G. de Vries, K. Correns y E. Cermak, publicaron de forma independiente los resultados de sus estudios, reproduciendo los experimentos de Mendel y confirmaron la exactitud de sus conclusiones. Dado que en ese momento se había descrito completamente la mitosis, casi por completo la meiosis (su descripción completa se completó en 1905), así como el proceso de fertilización, los científicos lograron conectar el comportamiento de los factores hereditarios mendelianos con el comportamiento de los cromosomas durante la célula. división. El redescubrimiento de las leyes de Mendel se convirtió en el punto de partida para el desarrollo de la genética.
La primera década del siglo XX. se convirtió en el período de la marcha triunfal del mendelismo. Los patrones descubiertos por Mendel fueron confirmados en el estudio de diversas características de objetos tanto vegetales como animales. Surgió la idea de la universalidad de las leyes de Mendel. Al mismo tiempo, comenzaron a acumularse hechos que no encajaban en el marco de estas leyes. Pero fue el método hibridológico el que permitió aclarar la naturaleza de estas desviaciones y confirmar la exactitud de las conclusiones de Mendel.
Todos los pares de caracteres que utilizó Mendel se heredaron según el tipo de dominancia completa. En este caso, el gen recesivo en el heterocigoto no tiene ningún efecto y el fenotipo del heterocigoto está determinado únicamente por el gen dominante. Sin embargo, una gran cantidad de rasgos en plantas y animales se heredan según el tipo de dominancia incompleta. En este caso, el híbrido F 1 no reproduce completamente el rasgo de uno u otro progenitor. La expresión del rasgo es intermedia, con mayor o menor desviación en una dirección u otra.
Un ejemplo de dominancia incompleta puede ser el color rosa intermedio de las flores en los híbridos de belleza nocturna obtenidos al cruzar plantas con un color rojo dominante y un blanco recesivo (ver diagrama).
Esquema de dominancia incompleta en la herencia del color de las flores en la belleza nocturna.

Como puede verse en el diagrama, al cruzar se aplica la ley de uniformidad de los híbridos de primera generación. Todos los híbridos tienen el mismo color (rosa) como resultado de la dominancia incompleta del gen. A. En la segunda generación, diferentes genotipos tienen la misma frecuencia que en el experimento de Mendel, y sólo cambia la fórmula de segregación fenotípica. Coincide con la fórmula de segregación por genotipo - 1: 2: 1, ya que cada genotipo tiene su propia característica. Esta circunstancia facilita el análisis, ya que no es necesario realizar cruces analíticos.
Existe otro tipo de comportamiento de los genes alélicos en un heterocigoto. Se llama codominancia y se describe en el estudio de la herencia de grupos sanguíneos en humanos y varios animales domésticos. En este caso, un híbrido cuyo genotipo contiene ambos genes alélicos exhibe ambos rasgos alternativos por igual. La codominancia se observa al heredar grupos sanguíneos del sistema A, B, 0 en humanos. personas con un grupo AB(grupo IV) hay dos antígenos diferentes en la sangre, cuya síntesis está controlada por dos genes alélicos.
La cuadrícula de Punnett es una herramienta visual que ayuda a los genetistas a determinar posibles combinaciones de genes durante la fertilización. Una cuadrícula de Punnett es una tabla simple de 2x2 (o más) celdas. Utilizando esta tabla y el conocimiento de los genotipos de ambos padres, los científicos pueden predecir qué combinaciones de genes son posibles en la descendencia e incluso determinar la probabilidad de heredar ciertos rasgos.
Pasos
Información básica y definiciones.
Para omitir esta sección e ir directamente a la descripción de la red de Punnett, .
- La reproducción sexual no es la única forma en que se reproducen los organismos vivos. Algunos organismos (por ejemplo, muchos tipos de bacterias) se reproducen a través de reproducción asexual, cuando la descendencia es creada por uno de los padres. En la reproducción asexual, todos los genes se heredan de uno de los padres y la descendencia es una copia casi exacta del mismo.
-
Conozca el concepto de alelos. Como se señaló anteriormente, los genes de un organismo vivo son un conjunto de instrucciones que le dicen a cada célula qué hacer. De hecho, al igual que las instrucciones ordinarias que se dividen en capítulos, cláusulas y subcláusulas independientes, las diferentes partes de los genes indican cómo hacer diferentes cosas. Si dos organismos tienen "subdivisiones" diferentes, se verán o se comportarán de manera diferente; por ejemplo, las diferencias genéticas pueden hacer que una persona tenga cabello oscuro y otra tenga cabello claro. Estos diferentes tipos de un gen se llaman alelos.
- Como el niño recibe dos conjuntos de genes, uno de cada padre, tendrá dos copias de cada alelo.
-
Conozca el concepto de alelos dominantes y recesivos. Los alelos no siempre tienen la misma "fuerza" genética. Algunos alelos que se llaman dominante, seguramente se manifestarán en la apariencia y el comportamiento del niño. Otros, los llamados recesivo Los alelos aparecen sólo si no coinciden con los alelos dominantes que los “suprimen”. La cuadrícula de Punnett se utiliza a menudo para determinar la probabilidad de que un niño reciba un alelo dominante o recesivo.
Representación de un cruce monohíbrido (un gen)
-
Dibuja una cuadrícula de 2x2. La versión más sencilla de la celosía de Punnett es muy fácil de hacer. Dibuja un cuadrado lo suficientemente grande y divídelo en cuatro cuadrados iguales. De esta forma tendrás una tabla con dos filas y dos columnas.
En cada fila y columna, etiquete los alelos padres con una letra. En una cuadrícula de Punnett, las columnas están reservadas para los alelos maternos y las filas para los alelos paternos, o viceversa. En cada fila y columna, escribe las letras que representan los alelos de la madre y del padre. Al hacerlo, utilice letras mayúsculas para los alelos dominantes y minúsculas para los recesivos.
- Esto es fácil de entender con un ejemplo. Suponga que desea determinar la probabilidad de que una pareja determinada tenga un hijo que pueda curvar la lengua. Esta propiedad se puede denotar en letras latinas. R Y r- una letra mayúscula corresponde a un alelo dominante y una letra minúscula corresponde a un alelo recesivo. Si ambos padres son heterocigotos (tienen una copia de cada alelo), entonces debes escribir una letra "R" y una "r" encima del hash Y una "R" y una "r" a la izquierda de la rejilla.
-
Escribe las letras correspondientes en cada celda. Puede completar fácilmente la cuadrícula de Punnett una vez que comprenda qué alelos están incluidos de cada padre. Escribe en cada celda una combinación de genes de dos letras que representen alelos de la madre y el padre. En otras palabras, tome las letras de la fila y columna correspondientes y escríbalas en la celda indicada.
Determinar los posibles genotipos de la descendencia. Cada celda de una cuadrícula de Punnett llena contiene un conjunto de genes que son posibles en un hijo de estos padres. Cada celda (es decir, cada conjunto de alelos) tiene la misma probabilidad; en otras palabras, en una red de 2x2, cada una de las cuatro opciones posibles tiene una probabilidad de 1/4. Las diferentes combinaciones de alelos representados en la cuadrícula de Punnett se denominan genotipos. Aunque los genotipos representan diferencias genéticas, esto no significa necesariamente que cada variante producirá una descendencia diferente (ver más abajo).
- En nuestro ejemplo de una cuadrícula de Punnett, un par determinado de padres puede producir los siguientes genotipos:
- Dos alelos dominantes(celda con dos R)
- (celda con una R y una r)
- Un alelo dominante y otro recesivo.(celda con R y r): tenga en cuenta que este genotipo está representado por dos celdas
- Dos alelos recesivos(celda con dos r)
-
Determinar los posibles fenotipos de la descendencia. fenotipo Los rasgos físicos reales de un organismo que se basan en su genotipo. Un ejemplo de fenotipo es el color de ojos, el color de cabello, la presencia de anemia falciforme, etc., aunque todos estos rasgos físicos están determinadas genes, ninguno de ellos está determinado por su combinación especial de genes. El posible fenotipo de la descendencia está determinado por las características de los genes. Diferentes genes se expresan de manera diferente en el fenotipo.
- Supongamos en nuestro ejemplo que el gen responsable de la capacidad de hacer girar la lengua es dominante. Esto significa que incluso aquellos descendientes cuyo genotipo incluya solo un alelo dominante podrán enrollar su lengua en un tubo. En este caso se obtienen los siguientes fenotipos posibles:
- Celda superior izquierda: puede curvar la lengua (dos R)
- Celda superior derecha:
- Celda inferior izquierda: puede rodar la lengua (una R)
- Celda inferior derecha: no puede rodar la lengua (sin R mayúsculas)
-
Determine la probabilidad de diferentes fenotipos en función del número de células. Uno de los usos más comunes de la cuadrícula de Punnett es encontrar la probabilidad de que un fenotipo determinado ocurra en la descendencia. Dado que cada célula corresponde a un genotipo específico y la probabilidad de aparición de cada genotipo es la misma, para encontrar la probabilidad de un fenotipo es suficiente dividir el número de células con un fenotipo dado por el número total de células.
- En nuestro ejemplo, la red de Punnett nos dice que para determinados padres, son posibles cuatro tipos de combinaciones de genes. Tres de ellos corresponden a un descendiente que es capaz de enrollar la lengua, y uno corresponde a la ausencia de tal habilidad. Así, las probabilidades de los dos fenotipos posibles son:
- El descendiente puede rodar la lengua: 3/4 = 0,75 = 75%
- El descendiente no puede rodar la lengua: 1/4 = 0,25 = 25%
Representación de un cruce dihíbrido (dos genes)
-
Divide cada celda de la cuadrícula de 2x2 en cuatro cuadrados más. No todas las combinaciones de genes son tan simples como el cruce monohíbrido (monogénico) descrito anteriormente. Algunos fenotipos están determinados por más de un gen. En tales casos, se deben tener en cuenta todas las combinaciones posibles, lo que requerirá oh mesa más grande.
- La regla básica para utilizar la cuadrícula de Punnett cuando hay más de un gen es la siguiente: Por cada gen adicional, se debe duplicar el número de células.. En otras palabras, para un gen es apropiada una cuadrícula de 2x2, para dos genes es apropiada una cuadrícula de 4x4, para tres genes es apropiada una cuadrícula de 8x8, y así sucesivamente.
- Para que sea más fácil comprender este principio, consideremos un ejemplo de dos genes. Para ello tendremos que dibujar una celosía. 4x4. El método descrito en esta sección también es adecuado para tres o más genes; sólo necesita oh Cuadrícula más grande y más trabajo.
-
Identificar genes del lado de los padres. El siguiente paso es encontrar los genes parentales responsables del rasgo que le interesa. Como se trata de múltiples genes, es necesario agregar una letra más al genotipo de cada padre; en otras palabras, es necesario utilizar cuatro letras para dos genes, seis letras para tres genes, y así sucesivamente. Como recordatorio, es útil escribir el genotipo de la madre encima de la almohadilla y el genotipo del padre a la izquierda (o viceversa).
-
Escribe las diferentes combinaciones de genes a lo largo de los bordes superior e izquierdo de la cuadrícula. Ahora podemos escribir arriba y a la izquierda de la cuadrícula los diversos alelos que cada padre puede transmitir a la descendencia. Como ocurre con un solo gen, cada alelo tiene la misma probabilidad de transmitirse. Sin embargo, dado que estamos analizando varios genes, cada fila o columna tendrá varias letras: dos letras para dos genes, tres letras para tres genes, y así sucesivamente.
- En nuestro caso, deberíamos anotar las distintas combinaciones de genes que cada padre es capaz de transmitir a partir de su genotipo. Si el genotipo de la madre es SsYy arriba y el genotipo SsYY del padre está a la izquierda, entonces para cada gen obtendremos los siguientes alelos:
- A lo largo del borde superior: SY, SY, SY, SY
- A lo largo del borde izquierdo: SY, SY, SY, SY
-
Complete las celdas con las combinaciones de alelos apropiadas. Escribe letras en cada celda de la cuadrícula de la misma manera que lo hiciste para un gen. Sin embargo, en este caso, por cada gen adicional, aparecerán dos letras adicionales en las celdas: en total, en cada celda habrá cuatro letras para dos genes, seis letras para cuatro genes, y así sucesivamente. Según la regla general, el número de letras de cada celda corresponde al número de letras del genotipo de uno de los padres.
- En nuestro ejemplo, las celdas se completarán de la siguiente manera:
- Fila superior: SSYY, SSYy, SsYY, SsYy
- Segunda fila: SSYY, SSYy, SsYY, SsYy
- Tercera fila: SsYY, SsYy, ssYY, ssYy
- Fila inferior: SsYY, SsYy, ssYY, ssYy
-
Encuentre los fenotipos para cada posible descendencia. En el caso de varios genes, cada célula en la red de Punnett también corresponde a un genotipo separado de posible descendencia; simplemente hay más de estos genotipos que con un solo gen. Y en este caso, los fenotipos de una célula en particular están determinados por los genes que estamos considerando. Existe una regla general que para que se manifiesten los rasgos dominantes es suficiente la presencia de al menos un alelo dominante, mientras que para los rasgos recesivos es necesario que Todo los alelos correspondientes eran recesivos.
- Dado que la suavidad y los granos amarillos son dominantes en los guisantes, en nuestro ejemplo, cualquier celda con al menos una S mayúscula corresponde a una planta con guisantes lisos, y cualquier celda con al menos una Y mayúscula corresponde a una planta con un fenotipo de grano amarillo. Las plantas con guisantes arrugados estarán representadas por células con dos alelos s minúsculas, pero solo se requieren ys minúsculas para que los granos sean verdes. Así, obtenemos posibles opciones para la forma y color de los guisantes:
- Fila superior:
- Segunda fila: liso/amarillo, liso/amarillo, liso/amarillo, liso/amarillo
- Tercera fila:
- Fila inferior: liso/amarillo, liso/amarillo, arrugado/amarillo, arrugado/amarillo
-
Determine la probabilidad célula por célula de cada fenotipo. Para encontrar la probabilidad de diferentes fenotipos en la descendencia de determinados padres, utilice el mismo método que para un solo gen. En otras palabras, la probabilidad de un fenotipo particular es igual al número de células correspondientes dividido por el número total de células.
- En nuestro ejemplo, la probabilidad de cada fenotipo es:
- Descendiente con guisantes lisos y amarillos: 12/16 = 3/4 = 0,75 = 75%
- Descendiente con guisantes arrugados y amarillos: 4/16 = 1/4 = 0,25 = 25%
- Descendiente con guisantes lisos y verdes: 0/16 = 0%
- Descendiente con guisantes arrugados y verdes: 0/16 = 0%
- Tenga en cuenta que la incapacidad de heredar los dos alelos recesivos y ha dado como resultado que no haya plantas con granos verdes entre la posible descendencia.
- Recuerde que cada nuevo gen parental duplica el número de células en la cuadrícula de Punnett. Por ejemplo, con un gen de cada padre se obtiene una cuadrícula de 2x2, con dos genes se obtiene una cuadrícula de 4x4, y así sucesivamente. En el caso de cinco genes, ¡el tamaño de la tabla será 32x32!
-
Aprenda más sobre el concepto de genes. Antes de comenzar a aprender y utilizar la cuadrícula de Punnett, debe familiarizarse con algunos principios y conceptos básicos. El primero de estos principios es que todos los seres vivos (desde los diminutos microbios hasta las gigantescas ballenas azules) tienen genes. Los genes son conjuntos microscópicos de instrucciones increíblemente complejos que están integrados en prácticamente todas las células de un organismo vivo. En esencia, en un grado u otro, los genes son responsables de todos los aspectos de la vida de un organismo, incluido su aspecto, cómo se comporta y mucho, mucho más.
Conoce más sobre el concepto de reproducción sexual. La mayoría (pero no todos) de los organismos vivos que conoce producen descendencia a través de reproducción sexual. Esto significa que los individuos femeninos y masculinos aportan sus propios genes y su descendencia hereda aproximadamente la mitad de los genes de cada padre. La cuadrícula de Punnett se utiliza para visualizar las diferentes combinaciones de genes de los padres.
Es bien sabido que la composición de las redes de Punnett se utiliza ampliamente para resolver problemas genéticos en genética mendeliana. La capacidad de construir correctamente una red de Punnett será útil para escolares y estudiantes en lecciones de biología. Pero los genetistas profesionales también utilizan estas habilidades en su trabajo. ¿Qué es una cuadrícula de Punnett?
La cuadrícula de Punnett es un método gráfico propuesto por el genetista británico Reginald Punnett en 1906, que demuestra visualmente todas las combinaciones posibles de diferentes tipos de gametos en cruces específicos o experimentos de reproducción (cada gameto es una combinación de un alelo materno y uno paterno para cada uno, gen estudiado en el cruce).
La cuadrícula de Punnett parece una tabla bidimensional, donde los gametos de un padre están escritos en la parte superior y los gametos del segundo padre están escritos verticalmente en la parte izquierda. Y en las celdas de la tabla en la intersección de filas y columnas, los genotipos de la descendencia se registran como combinaciones de estos gametos. Esto hace que sea muy fácil determinar las probabilidades de cada genotipo en un cruce determinado.
Compilando una grilla de Punnett en un cruce monohíbrido
En un cruce monohíbrido, se examina la herencia de un gen. En un cruce monohíbrido clásico, cada gen tiene dos alelos. Por ejemplo, tomaremos organismos maternos y paternos con el mismo genotipo: "Gg". En genética, las letras mayúsculas se utilizan para indicar un alelo dominante y las minúsculas para un alelo recesivo. Este genotipo puede producir sólo dos tipos de gametos, que contienen el alelo "G" o el alelo "g".
Nuestra red Punnett se verá así:
| GRAMO | gramo | |
| GRAMO | GG | gg |
| gramo | gg | ggg |
Al resumir los mismos genotipos en la red de Punnett para nuestra descendencia, obtenemos la siguiente proporción de genotipo: 1 (25%) GG: 2 (50%) GG: 1 (25%) GG es una proporción de genotipo típica (1:02 :01) para un cruce monohíbrido. El alelo dominante enmascarará el alelo recesivo, lo que significa que los organismos con genotipos "GG" y "Gg" tienen el mismo fenotipo.
Por ejemplo, si el alelo "G" produce un color amarillo y el alelo "g" produce un color verde, entonces el genotipo "gg" tendrá un fenotipo verde y los genotipos "GG" y "Gg" tendrán un fenotipo amarillo. fenotipo. Al resumir los valores en la cuadrícula, tendremos 3G- (fenotipo amarillo) y 1gg (fenotipo verde); esta es una proporción fenotípica típica (3:1) para un cruce monohíbrido. Y las probabilidades correspondientes para la descendencia serán 75%G-: 25%gg.
Red de Punnett y herencia mendeliana
Estos resultados se obtuvieron por primera vez en experimentos de Gregor Mendel con una planta: el guisante (Pisum sativum). Al interpretar los resultados, Mendel llegó a las siguientes conclusiones:
- Cada rasgo de un organismo determinado está controlado por un par de alelos.
- Si un organismo contiene dos alelos diferentes para un rasgo determinado, entonces uno de ellos (dominante) puede manifestarse, suprimiendo por completo la manifestación del otro (recesivo).
- Durante la meiosis, cada par de alelos se separa (divide) y cada gameto recibe uno de cada par de alelos (principio de escisión).
Sin estas leyes básicas no podremos resolver ningún problema genético. Habiendo establecido la capacidad de predecir resultados a partir de un par de rasgos alternativos, Mendel pasó a estudiar la herencia de dos pares de dichos rasgos.
Compilando una grilla de Punnett en un cruce dihíbrido
Los cruces dihíbridos examinan la herencia de dos genes. Para los cruces dihíbridos, podemos hacer una red de Punnett solo si los genes se heredan independientemente entre sí; esto significa que cuando se forman los gametos maternos y paternos, cualquier alelo de un par puede ingresar a cada uno de ellos junto con cualquier otro de otro par. . Este principio de distribución independiente fue descubierto por Mendel en experimentos con cruces dihíbridos y polihíbridos.
Tenemos dos genes: forma y color. Para la forma: "R" es el alelo dominante, que da una forma de guisante suave, y "w" es el alelo recesivo, que da una forma de guisante arrugada. Para el color: "Y" es el alelo dominante, que determina el color amarillo, y "g" es el alelo recesivo, que da el color verde de los guisantes. Las plantas masculinas y femeninas tienen el mismo genotipo: "RwYg" (liso, amarillo).
Primero necesitas determinar todas las combinaciones posibles de gametos, para esto también puedes usar la cuadrícula de Punnett:
| R | w | |
| R | R.R. | rw |
| w | rw | ww |
Así, las plantas heterocigotas pueden producir cuatro tipos de gametos con todas las combinaciones posibles: RY, Rg, wY, wg. Ahora creemos una red de Punnett para genotipos:
| RY | rg | wY | wg | |
| RY | RRHH | RRYg | RwYY | RwYg |
| rg | RRYg | RRgg | RwYg | rwgg |
| wY | RwYY | RwYg | wwYY | wwYg |
| wg | RwYg | rwgg | wwYg | wwgg |
Sumando los mismos genotipos en la red de Punnett para nuestra descendencia, obtenemos la siguiente proporción y probabilidades por genotipo: 1(6,25%) RRYY: 2(12,5%) RwYY: 1(6,25%) wwYY: 2(12,5%) RRYg: 4(25%) RwYg: 2(12,5%) wwYg: 1(6,25%) RRgg: 2(12,5%) Rwgg: 1(6,25%) wwgg. Y dado que los rasgos dominantes enmascaran los recesivos, obtenemos la siguiente proporción y probabilidades para los fenotipos: 9(56,25%) R-Y- (liso, amarillo): 3(18,75%) R-gg (liso, verde): 3 (18,75%) wwY- (arrugado, amarillo): 1(6,25%)wwgg (arrugado, verde). Esta proporción fenotípica, 9:3:3:1, es típica de un cruce dihíbrido.
Construcción de una rejilla de Punnett en un cruce trihíbrido.
Construir una cuadrícula de Punnett para un cruce entre dos plantas heterocigotas para tres genes será más difícil. Para resolver este problema podemos utilizar nuestros conocimientos de matemáticas. Para determinar todas las combinaciones posibles de gametos para un cruce trihíbrido, debemos recordar la solución de polinomios.
- Hagamos un polinomio para este cruce: (A + a) X (B + b) X (C + c).
- Multipliquemos la expresión del primer paréntesis por la expresión del segundo; obtenemos: (AB + Ab + aB + ab) X (C + c).
- Ahora multiplique esta expresión por la expresión del tercer paréntesis; obtenemos: ABC + ABc + AbC + Abc + aBC + aBc + abC + abc.
En consecuencia, pueden producir ocho tipos de gametos con todas las combinaciones posibles. Esta solución se puede ilustrar utilizando una red de Punnett:
| A | a | |
| B | AB | AB |
| b | ab | ab |
| C | C | |
| AB | A B C | A B C |
| ab | A B C | A B C |
| AB | a B C | a B C |
| ab | a B C | a B C |
Ahora creemos una red de Punnett para genotipos (la tabla tendrá 64 celdas):
| A B C | a B C | A B C | a B C | A B C | a B C | A B C | a B C | |
| A B C | AABBCC | AaBBCC | AABbCC | AaBbCC | AABBcc | AaBBCc | AABbCc | AaBbCc |
| a B C | AaBBCC | aaBBCC | AaBbCC | aaBbCC | AaBBCc | aaBBBCc | AaBbCc | aaBbCc |
| A B C | AABbCC | AaBbCC | AAbbCC | AabbCC | AABbCc | AaBbCc | AAbbCc | AabbCc |
| a B C | AaBbCC | aaBbCC | AabbCC | aabbCC | AaBbCc | aaBbCc | AabbCc | aabbCc |
| A B C | AABBcc | AaBBCc | AABbCc | AaBbCc | AABBcc | AaBBcc | AABbcc | AaBbcc |
| a B C | AaBBCc | aaBBBCc | AaBbCc | aaBbCc | AaBBcc | aaBBcc | AaBbcc | aaBbcc |
| A B C | AABbCc | AaBbCc | AAbbCc | AabbCc | AABbcc | AaBbcc | AAbbcc | Aabbcc |
| a B C | AaBbCc | aaBbCc | AabbCc | aabbCc | AaBbcc | aaBbcc | Aabbcc | aabbcc |
Los individuos de la generación F 1 son el resultado del cruce de dos organismos progenitores: masculino y femenino. Cada uno de ellos puede formar una determinada cantidad de tipos de gametos. Cada gameto de un organismo tiene la misma probabilidad de encontrarse con cualquier gameto de otro organismo durante la fertilización. Por tanto, el número total de cigotos posibles se puede calcular multiplicando todos los tipos de gametos de ambos organismos.
Cruz monohíbrida
Ejemplo 7.1. Anote el genotipo de los individuos de la primera generación al cruzar dos individuos: homocigotos para un gen dominante y homocigotos para un gen recesivo.
Escribamos la letra que designa los genotipos de los pares parentales y los gametos que forman.
R AA x aa
Gametos A
En este caso, cada uno de los organismos produce gametos del mismo tipo, por lo tanto, cuando los gametos se fusionan siempre se formarán individuos con el genotipo Aa. Los individuos híbridos desarrollados a partir de tales gametos serán uniformes no sólo en genotipo, sino también en fenotipo: todos los individuos portarán un rasgo dominante (según la primera ley de uniformidad de la primera generación de Mendel).
Para que sea más fácil registrar los genotipos de la descendencia, el encuentro de los gametos suele indicarse mediante una flecha o una línea recta que conecta los gametos del organismo masculino y femenino.
Ejemplo 7.2. Determinar y registrar los genotipos de individuos de primera generación al cruzar dos individuos heterocigotos analizados para el mismo rasgo.
R Aa x Aa
Gametos A; una A; A ![]()
F1AA; Aa Aa; ah
Cada padre produce dos tipos de gametos. Las flechas indican que cualquiera de los dos gametos de un individuo femenino puede encontrarse con cualquiera de los dos gametos de un individuo masculino. Por tanto, son posibles cuatro variantes de gametos y en la descendencia se forman individuos con los siguientes genotipos: AA, Aa, Aa, aa.
Ejemplo 7.3. El cabello puede ser de color claro u oscuro. El gen del color oscuro es dominante. Una mujer heterocigota y un hombre homocigoto de cabello oscuro contrajeron matrimonio. ¿Qué genotipos se deben esperar en los niños de la primera generación?
rasgo: gen
color oscuro: A
color claro: un
R AA x AA
oscuro oscuro
Gametos A; una una
oscuro oscuro
Cruz dihíbrida
El número y los tipos de cigotos en un cruce dihíbrido dependen de cómo están dispuestos los genes no alélicos.
Si en un par de cromosomas homólogos se ubican genes no alélicos responsables de diferentes rasgos, entonces el número de tipos de gametos en un organismo diheterocigoto con genotipo Aa BB será igual a dos: AB y ab. Cuando se cruzan dos de estos organismos, la fertilización dará como resultado la formación de cuatro cigotos. La grabación de los resultados de dicho cruce se verá así:
R AVav x Avav
Gametos AB; av av; ay
F1ABAB; ABav; ABav; avaw
Los organismos diheterocigotos que contienen genes no alélicos en cromosomas no homólogos tienen el genotipo AaBb y forman cuatro tipos de gametos.
Al cruzar dos de estos individuos, las combinaciones de sus gametos darán 4x4 = 16 opciones de genotipo. El genotipo de los individuos resultantes se puede registrar secuencialmente uno tras otro, como hicimos con el cruce monohíbrido. Sin embargo, tal registro línea por línea será demasiado engorroso y difícil para el análisis posterior. El genetista inglés Punnett propuso registrar el resultado del cruce en forma de una tabla que lleva el nombre del científico: la cuadrícula de Punnett.
Primero, se registran como de costumbre los genotipos de las parejas parentales y los tipos de sus gametos, luego se dibuja una cuadrícula en la que el número de columnas verticales y horizontales corresponde al número de tipos de gametos de los individuos parentales. Los gametos del individuo femenino están escritos horizontalmente en la parte superior y los gametos del individuo masculino están escritos verticalmente a la izquierda. En la intersección de líneas imaginarias verticales y horizontales provenientes de los gametos de los padres, se registran los genotipos de la descendencia.